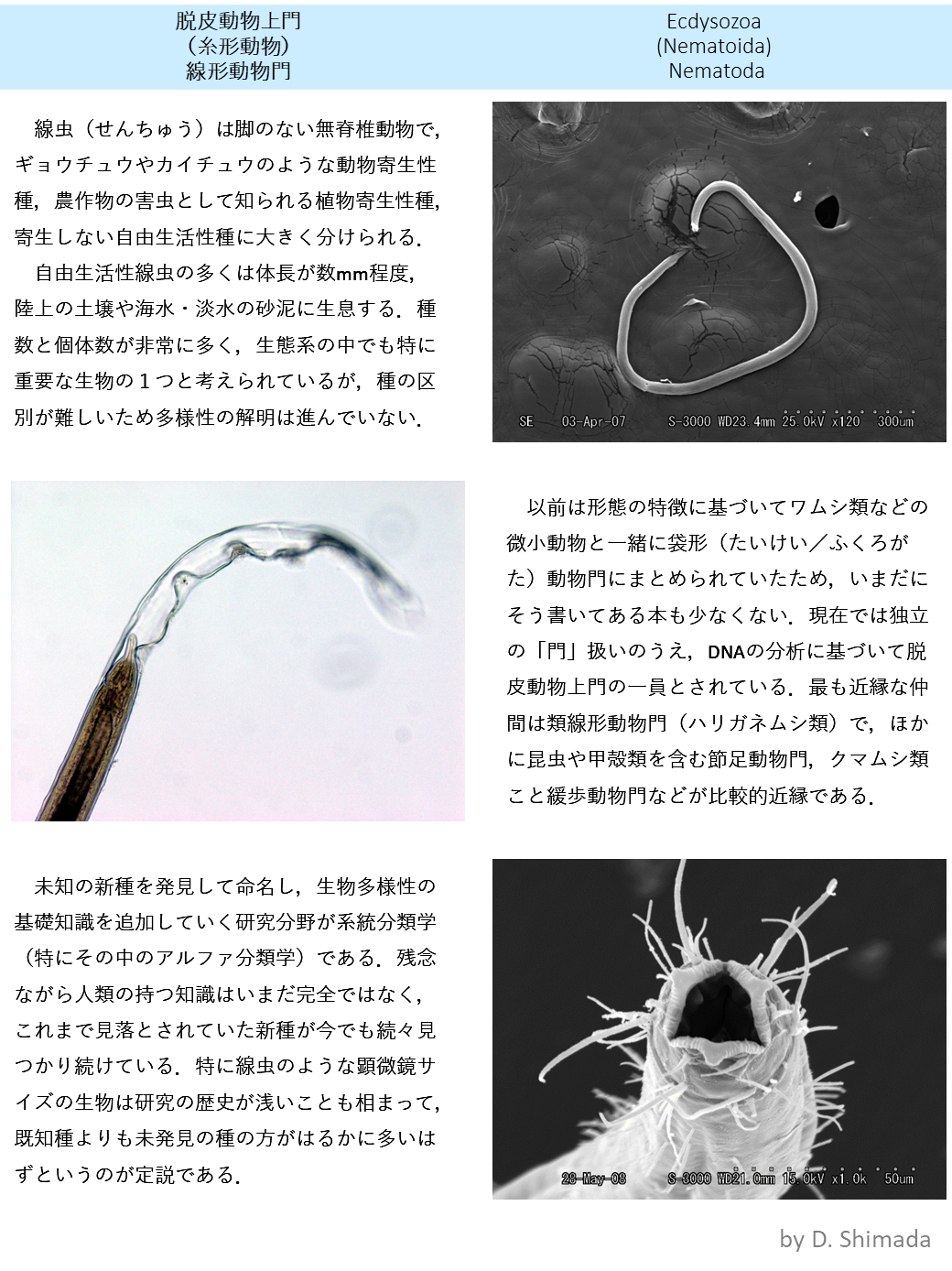
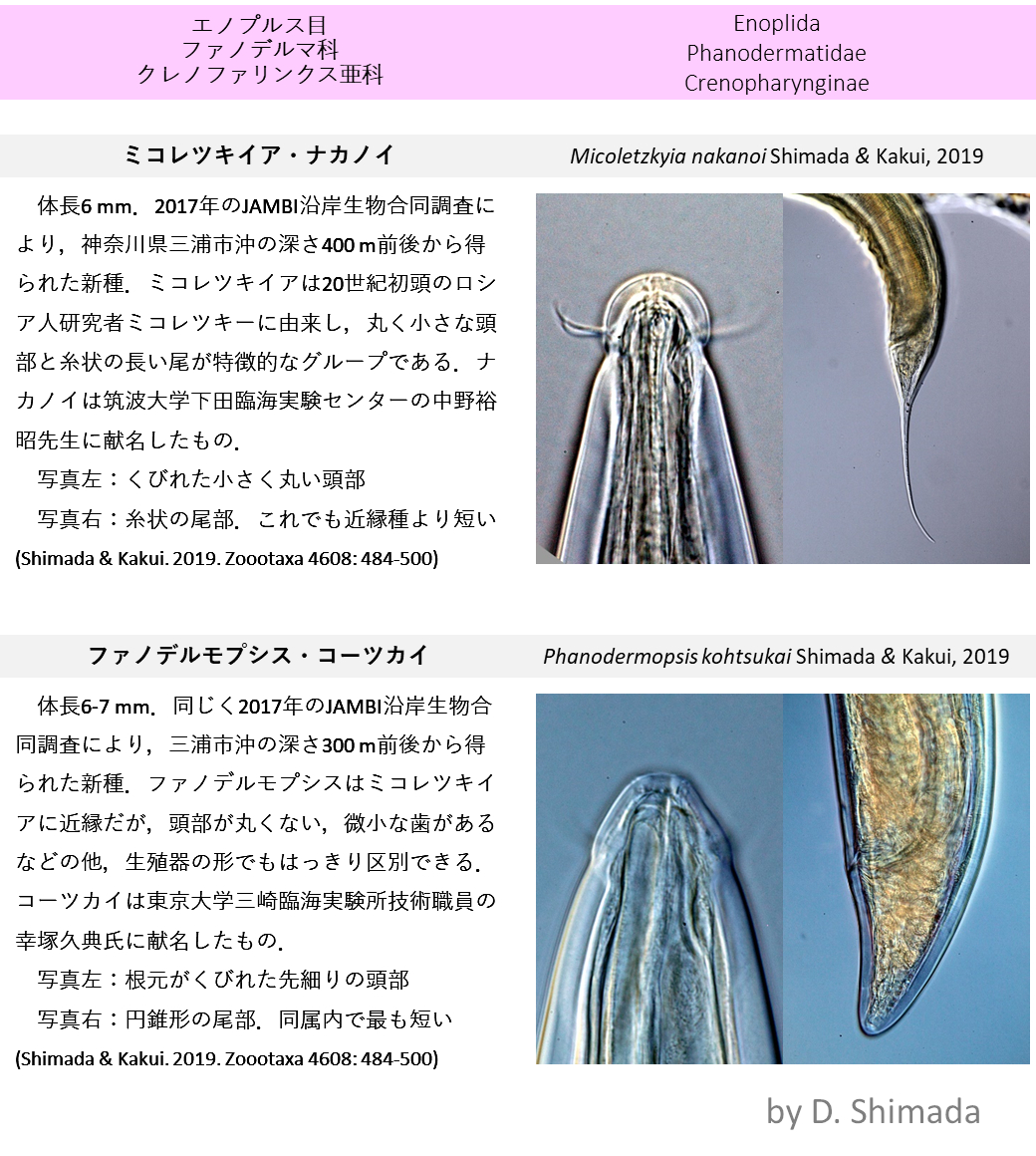
ギャラリー

















生理条件下で上皮細胞の残渣を貪食する間充織細胞
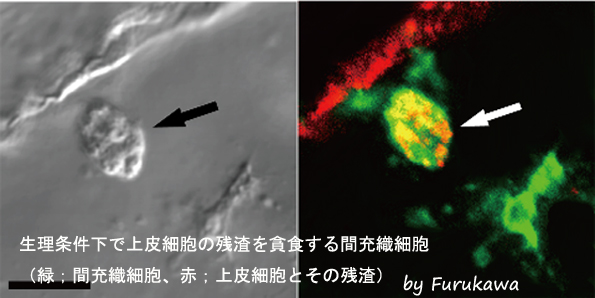
注射によって人為的に導入した異物に対して、ヒトデ幼生の間充織細胞が免疫応答を示すことは古くから知られていた。しかし、実際に生理条件下で、間充織細胞が生体防御を行う細胞であるかについては明らかになっていなかった。発生過程で生じた死細胞などは、生体にとって内因性の異物と見なされ、免疫細胞によって貪食される。この現象を指標として、上皮細胞マーカー及び間充織細胞マーカーを用いて、間充織細胞が死んだ上皮細胞を生理条件下で貪食しているか解析した。その結果、緑でラベルされた間充織細胞(矢印)が、赤でラベルされた上皮細胞の残渣を貪食している様子が観察され、間充織細胞が生理条件下でも免疫細胞として機能していることが明らかとなった。
水晶中の原子配列 赤が酸素、青がケイ素
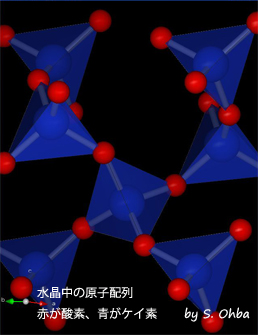
一般に組成式がSiO2で表わせるものをシリカといい,ガラスの状態もとりうるが,結晶相も多数知られている。その1つが石英であり,石英の結晶面がきれいに発達したものを特に水晶とよんでいる。通常,水晶は六角柱状で先がとがっており,その伸長方向が結晶のc軸であり,これが旋光性を示す光軸ともなっている。石英の結晶中のSiO4の四面体のつながり方に注目すると,結晶の伸長方向(c軸)に対してラセンを形成しており,このラセンの巻き方が右巻きの結晶と左巻きの結晶とが存在する。α-石英の結晶構造の絶対配置はX線の異常散乱を用いて決定された(1958年)。これにより,右旋性石英中の四面体の配列が左巻きラセンであることが判明した。さらに理論的な検討により,他の化合物も含めて,旋光性に関与するキラルな原子配列が右巻きラセンのときは左旋性,原子配列が左巻きラセンのときは右旋性となることが示された(1986年)。
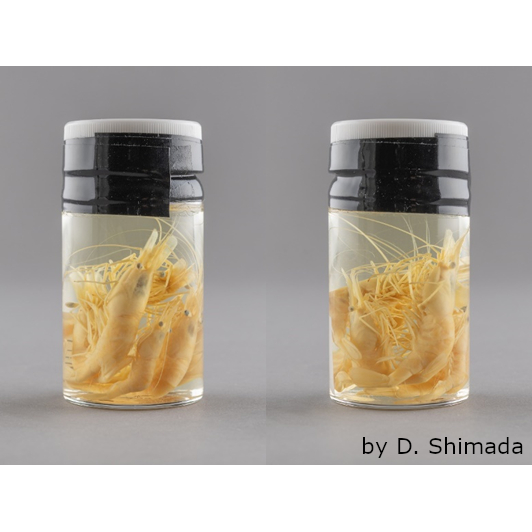
イトマキヒトデ(
Asterina pectinifera)幼稚体
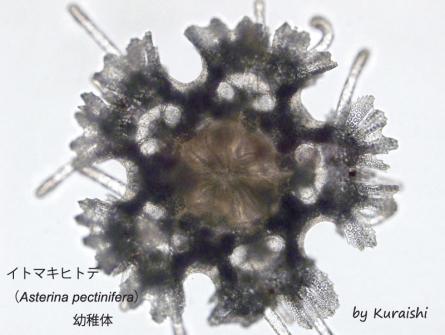
多くのヒトデ類の幼性は数週間の幼生期間を経た後に基質(モミジガイの場合は砂、イトマキヒトデの場合は海藻や岩など)に付着して変態する。この時、幼生の体の前の部分は退縮してしまい、その後部だけが「ヒトデ」の体になる。そのため、変態直前の幼生の体長は1mmを超えるのに対し、変態直後の幼稚体の直径は500µmほどしか無い。
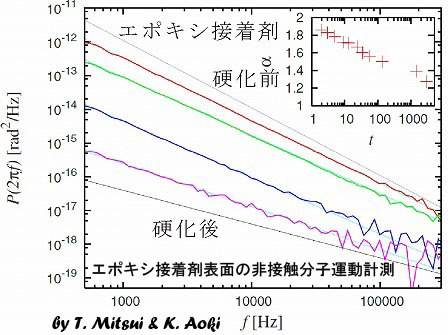
アナログ集積回路のICプローブによる測定。赤は電力を多く消費している部分

半導体集積回路(IC)の正常動作状態における素子内部の電流分布を測定する実験。半導体素子内部に電流が流れると発熱し、熱膨張長するので、集積回路の表面形状変化を測定することにより、電流分布がわかる。図は正弦波の増幅を行っている演算増幅器の測定例である。赤で示した部分が正弦波の周期で熱膨張を繰り返している部分。

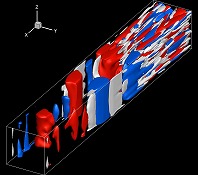
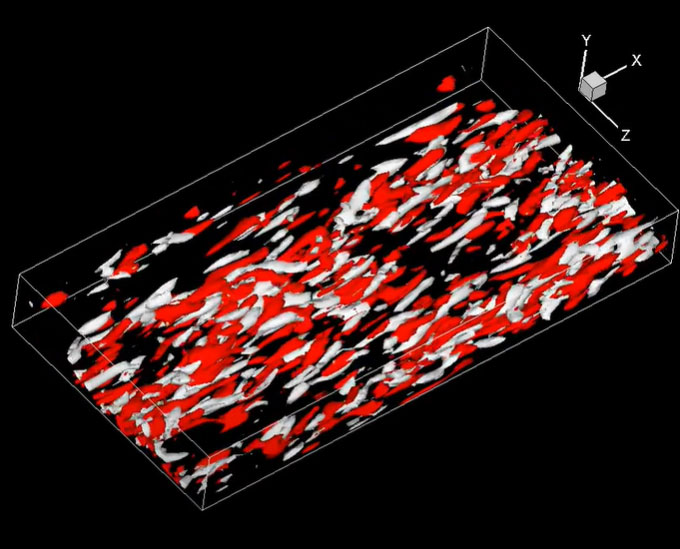
局所磁場を印加した液体金属ダクト流れの乱流構造
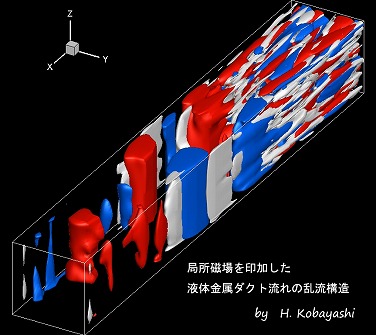
車や飛行機の周りの空気の流れなど,身の回りには乱れた流れである乱流が存在する。その乱流中では大きなスケールの渦から小さなスケールの渦へ分裂していき,熱になって最後は散逸する過程が含まれている。上図では四方を壁で囲まれた矩形ダクト内を水銀などに代表される液体金属流体が右上から乱流状態で流入している様子を描画している。流路中央部分には下から上に向かって磁場が印可されている。白い領域が強い回転渦が存在する領域で,赤と青の領域はそれぞれレイノルズ応力が正・負となる領域を示している。乱流状態の液体金属が磁場中を通過するとその領域で乱流が抑制され,磁場の方向に渦やレイノルズ応力などの乱流構造が揃う様子が分かる。
オニクマムシ
Milnesium tardigradumの親(右)とその脱皮殻の中の卵(左)
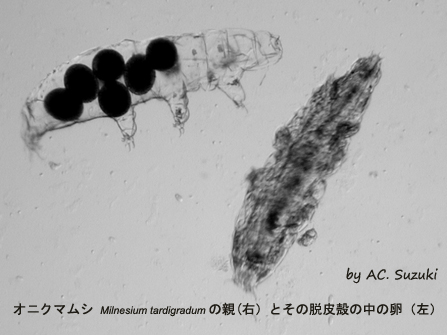
オニクマムシは2回脱皮して、3齢となって性成熟し、3回目の脱皮からは産卵が伴う。脱皮の前に、新旧のクチクラ(体表の硬い殻の部分)の隙き間に産卵し、その後で古いクチクラを脱ぎ捨てる。その結果、卵は古いクチクラの中で保護された状態となる。
オニクマムシ
Milnesium tardigradumは脱皮する前に 古いクチクラの中で産卵する
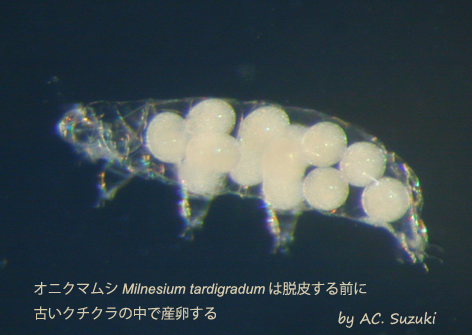
オニクマムシは一度に1~18個の卵を産む。その数は、親の栄養状態による。上の写真では、脱皮殻の中に13個の卵が見える。クマムシの種によっては、脱皮殻の中へ産卵するのではなく、普通に体の外へ産みつける様式のものもある。そのような種では、卵の表面に様々な突起が形成される場合が多く、そのような突起が卵を動きにくくする機能を持つと考えられる。しかしオニクマムシのように脱皮殻の中に産まれる卵の表面はつるつるで、目立った突起などはみられない。
ゆで卵を含む任意の高速回転軸対称物体の運動方程式

コロンブスは知っていたのだろうか、ゆで卵をテーブルに置いて速く回すと立ち上がる。碁石やラグビーボール等でも同じようなことが起きる。これらは良く知られた運動であるが、重心が重力に抗して上昇する点が不思議である。テーブルと回転物体との接点で働く摩擦力が不可欠で、系の力学的全エネルギーは運動につれて散逸していくけれど、回転が速い場合に断熱不変量が存在する。その結果、任意の軸対称剛体が満たす一階の常微分方程式が導出され、立ち上がりを示す解が得られる。
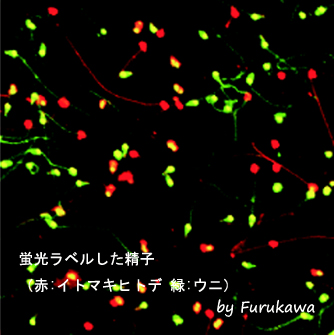
間充織細胞の移植実験
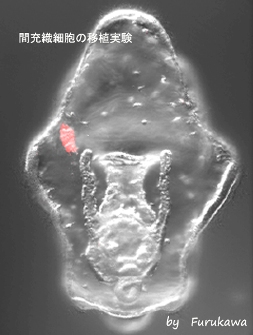
培養下に単離した間充織細胞を蛍光色素(赤)でラベルし、パラホルムアルデヒドで個体した状態又は生きたままの状態で、ヒトデ幼生に移植した。この移植された間充織細胞に対するホスト個体の免疫応答を観察することにより、ヒトデの幼生が同種異個体を識別する能力を有しているか確かめることができる。


棘皮動物イトマキヒトデ
Asterina pectinifera

約1800種とも言われる現存するヒトデ種の中で、日本近海で棲息する1種。イトマキヒトデ成体の体長は約5〜10㎝程度であり、漁港のテトラポッドや磯場で良く見かけられる。イトマキヒトデは雌雄異体であり、生殖シーズンになると5腕内にぎっしりと卵巣や精巣を含む。ウニと異なり、これらの生殖巣は食用にならないが、発生学の研究には有用な動物として注目されてきた。私たちの研究室では、人工海水中でイトマキヒトデ成体を飼育しながら、必要に応じて各生殖巣を成体から取り出し、1メチルアデニンと呼ばれる化学物質を用いて、卵を成熟させ、雄の精子を加えて、自在に発生を開始させることができる。外国の友人にイトマキヒトデ成体の実物を見せたところ、サイケデリックな色合が美しいとの感想であった。



イトマキヒトデ(
Asterina pectinifera) 初期原腸胚
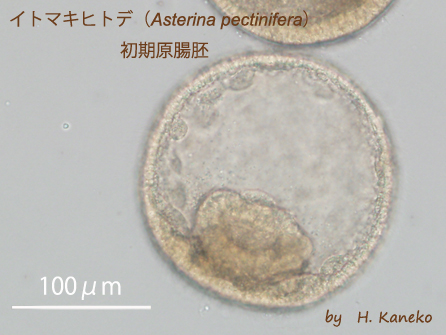
イトマキヒトデの胚期は、比較的速いスピードで発生が進行していく。画像は、20℃の人工海水中で発生させている受精18時間後の初期原腸胚である。球形の胚体の一部に、円筒を押しつぶした様な形が見られるが、これは2時間前に陥入し始めた原腸である。原腸は、胚体外部の外胚葉と切り離されておらず、連続した上皮構造である。胚体を構成する上皮構造が連続性を保ちながら、その一部がこの様に迅速、かつ顕著な形態変化を生じさせるメカニズムは、多くの発生学の研究者が興味を持ってきた。現在、原腸を構成する細胞群が個々に形や接着性を変えることが一因となっているらしいことが判っている。なお、原腸は、以後時間経過に伴って、胚体内を陥入していき、将来の消化管や体腔嚢を形成するようになる。
一様等方性乱流中の渦構造

車や飛行機の周りの空気の流れなど,身の回りには乱れた流れである乱流が存在する。その乱流中では大きなスケールの渦から小さなスケールの渦へ分裂していき,熱になって最後は散逸する過程が含まれている。上図では壁から遠く離れた場所に存在する一様等方な場での乱流中での渦構造を描画している。緑の領域が強い回転渦が存在する領域を示しており,渦の伸張方向はランダムである。多くの渦が集まっている領域(クラスター領域)と渦のない領域が存在することも分かる。
水晶(ハーキマーダイヤモンド)

高温の状態でβ-石英として結晶化したものは,温度が下がっても結晶外形はそのまま保たれるが内部構造はα-石英になる。これを高温形低温水晶とよぶ。通常の水晶は何か核となるところから放射状に成長するので,六角柱の片側だけが尖っている。しかし,Herkimer(ハーキマー)ダイヤモンドと呼ばれ,アメリカのニューヨーク州で産出する,やや小さめの水晶はころっとしていて,両端が尖っている。結晶面はすべて平坦であり,どこかに付着して成長したような跡はない。これは火山活動に伴い,二酸化ケイ素SiO2を含む高温の熱水の泡がドロマイトという堆積岩の隙間に入り,結晶が浮いた状態で1粒ずつ成長したためと推定される。結晶外形は基本的に六角柱の部分が短い両錐形である。この写真の例では(全長約7 mmの結晶の実体顕微鏡写真),正面に非対称な7角形の柱面(m)があり,その右上と左上および左下に微小面sが細長い長方形として現われている。もしα-石英(三方晶系)で双晶でないならば,微小面sと等価な位置は六角柱の上3ヶ所(および下3ヶ所)に限られる。しかし,ハーキマーダイヤモンドの結晶外形はこれに当てはまらないものが多くみられることから,結晶が生成したときはβ-石英の構造(六方晶系)であったと推定される。結晶化した後に温度が下がり,β形(高温形)からα形に転移したが,結晶外形は保存されている。このように,結晶構造の対称性と異なる外形をもつことを仮像といい,他の鉱物でも見られる現象である。
f 平面浅水系における非線形数値実験による流れ場のスナップショット
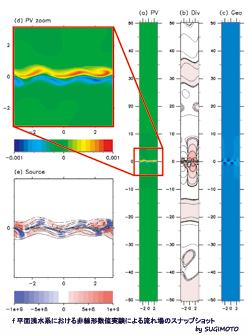
ジェットの蛇行から生じる大気重力波の数値実験(f平面浅水系)
大気や海洋などの地球の大規模な流れは、地球の自転と密度の成層の影響を大きく受ける。本研究では、それぞれの効果を含めて地球の流れを簡単に表現した、f平面浅水系を用いて、ジェット気流の蛇行(左上図:ポテンシャル渦度場)とそれに伴う大気重力波の放射過程(右図中:発散場)を、数値計算により調べている。左下図は渦からの音波放射理論(ライトヒル理論)を援用して導出した重力波のソース(カラー)であり、渦の領域(コンター)に集中していることがわかる。
大気や海洋などの地球の大規模な流れは、地球の自転と密度の成層の影響を大きく受ける。本研究では、それぞれの効果を含めて地球の流れを簡単に表現した、f平面浅水系を用いて、ジェット気流の蛇行(左上図:ポテンシャル渦度場)とそれに伴う大気重力波の放射過程(右図中:発散場)を、数値計算により調べている。左下図は渦からの音波放射理論(ライトヒル理論)を援用して導出した重力波のソース(カラー)であり、渦の領域(コンター)に集中していることがわかる。

培養間充織細胞の多核凝集体
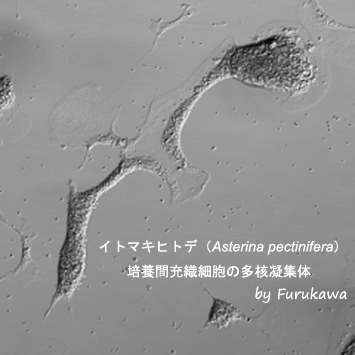
培養下に単離した間充織細胞を、培養基質から剥がして回収し、再度シャーレに播種すると、間充織細胞は速やかに図のような多核凝集体を形成する。この現象は、間充織細胞が培養基質を異物として認識し、免疫応答を惹起した結果ではないかと推測されている。一方、この状態を放置すると、間充織細胞は個々の細胞に分離し、再度ネットワーク様の構造を形成する。このような間充織細胞による融合と分離が、どのようなメカニズムで制御されているのか似関しては、全くわかっていない。
イトマキヒトデ (
Asterina pectinifera) 中期原腸胚 間充織細胞
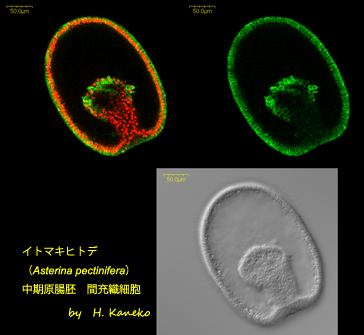
イトマキヒトデ胚の体制はかなりシンプルである。下図に見られるように、単層の上皮シートが、胚体の後方部で身体の内側に陥入して、将来の消化管となる原腸を作る。陥入位置となる原腸部は、将来の肛門となる。一方、胚体の内部まで陥入している原腸の先端は膨らんでおり、ここから体腔嚢と呼ばれる将来の排出器官が形成されていく。また、原腸先端部からは、単層上皮シートを解消して、個々の細胞として、胚体内部を活発に移動する間充織細胞が出現する。上画像において、緑色で標識された細胞が間充織細胞であり、将来的にはマクロファージ様の性質を帯び、幼生期の生体防御を担うようになる。
CMOS集積回路のICプローブによる測定
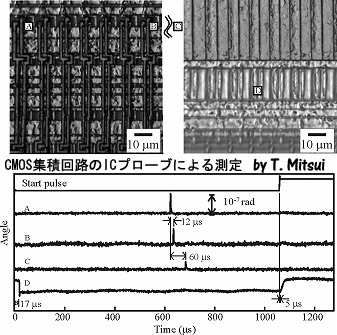
半導体集積回路(IC)の正常動作状態における素子内部の電流分布を測定する実験。半導体素子内部に電流が流れると発熱し、熱膨張長するので、集積回路の表面形状変化を測定することにより、電流分布がわかる。図はCMOSリニアイメージセンサの測定例である。
培養下に単離したヒトデ胚の間充織細胞

間充織細胞は、ヒトデ胚・幼生期における免疫細胞である。原腸先端から間充織細胞が出現し始めた中期原腸胚を、単一の細胞に解離しシャーレ上に播種すると、基質に対する接着能力を有する間充織細胞を、単一の細胞集団として図のように単離することができる。培養基質に接着した間充織細胞は、高度に仮足を発達させながらお互いに接触しあい、融合と分離を繰り返しながら図のようなネットワーク様の構造を形成する。生体内においても、間充織細胞によるこのようなネットワーク構造は局所的に形成されている。
水晶球のエアリースパイラル(2重渦巻き模様)

水晶球が左か右水晶かを区別するために,簡易型の偏光器(きらりビュアー)を用いる。偏光板を通過した光を水晶球に入れ,それをさらに円偏光板を通過させてから目で観察するようになっている。水晶球の光軸(c軸)がちょうど光の通過方向と一致したときだけ,虹色のエアリースパイラル(Airy’s spiral)が見える。そして,左水晶では左巻き,右水晶では右巻きの渦になるので,その渦巻きの方向から水晶球の右と左が区別できる。G. B. Airy(エアリー)はイギリスの天文学者であり,1836年から1881年にかけてグリニッジ天文台長にもなった人である。天文学以外の分野でも,結晶による干渉縞を調べ色消し接眼レンズの研究などを行った。エアリースパイラルは彼の名前に由来している。
平行平板間チャネル乱流中の渦構造

車や飛行機の周りの空気の流れなど,身の回りには乱れた流れである乱流が存在する。その乱流中では大きなスケールの渦から小さなスケールの渦へ分裂していき,熱になって最後は散逸する過程が含まれている。上図では上下におかれた平行平板間を流体が乱流状態で右下から左上へ流れている様を描画している。白い領域が強い回転渦が存在する領域で,緑の領域は乱流エネルギーが大きいスケールから小さいスケールへ散逸する領域を示している。強い渦の周りで乱流のエネルギー散逸が起こっており,そのような乱流構造は流れ方向に伸張していることも分かる。
ホスト個体内で共存する移植された間充織細胞

赤色蛍光物質でラベルしたイトマキヒトデの間充織細胞を、幼生の体内に生きた状態で注射した。図は移植24時間後の様子を示している。バクテリアやラテックスビーズなどを注射した場合、幼生の間充織細胞は注射された異物に対して速やかに応答し、2時間以内に包囲化作用や貪食作用といった攻撃を仕掛けるが、同種の間充織細胞は、24時間後においてもそのような攻撃を受けることなく、ホスト個体内で共存した。この事実は、ヒトデ幼生の免疫システムは、同種異個体の間充織細胞を異物と見なさないことを示唆している。
水晶のエアリースパイラル(旋光性による渦巻き模様)

この写真は,右水晶板(右水晶のc軸に垂直に切った板)の上に左水晶板(両方とも厚さ4 mm)をのせ,垂直配置の直線偏光板2枚ではさんで,カメラのレンズを接近させて撮影したものである。右回りの4重渦巻き模様となっている。これがエアリースパイラルの原型である。水晶板が正確にc軸に垂直でないと,干渉像はまだら模様となってしまうが,結晶板の方位を調整して,2枚の水晶板の干渉像の中心円がうまく重なるように合わせてやれば,4重渦巻きが見える。2枚の水晶板の厚さが多少違っていても問題はない。このように,原理的には右水晶と左水晶板を重ねて光軸の方向を一致させ,それに光を通過させなければならない。しかし,鏡による光の反射を利用すると,水晶板1枚だけで同様な観察をすることができる。水晶板が右結晶とすると,右水晶を通過して鏡で反射した光は,同じ厚さの左水晶板を通過した光と同等なので,もう一度右水晶板を通過させてから検光子を通過させる仕組がかつて用いられていた。この装置を使うと,左結晶と右結晶の区別の他に,切り出された水晶板の方位が正確にc軸に対して垂直かを検査することができる。
プラナリア人為的有性化個体とその複合卵

生物学的な意味で「性」とは、2個体のゲノムの混合を意味する。
再生能が高いことで有名なプラナリアは、切断と再生を繰り返し無性的な生殖で次世代を産生するのみならず、卵と精子が受精してゲノムを混合し、次世代の仔虫を含む複合卵を産卵する有性的な生殖も可能である。自然界の状況でこの2つの方法を切り換えているものもいる。如何にしてプラナリアは生殖方法を切り換えるのだろうか?私達は人為的に無性生殖をしているプラナリアを有性生殖に転換させる方法を開発した。この方法を用いてこのメカニズムを解明することを私達はめざしている。
再生能が高いことで有名なプラナリアは、切断と再生を繰り返し無性的な生殖で次世代を産生するのみならず、卵と精子が受精してゲノムを混合し、次世代の仔虫を含む複合卵を産卵する有性的な生殖も可能である。自然界の状況でこの2つの方法を切り換えているものもいる。如何にしてプラナリアは生殖方法を切り換えるのだろうか?私達は人為的に無性生殖をしているプラナリアを有性生殖に転換させる方法を開発した。この方法を用いてこのメカニズムを解明することを私達はめざしている。

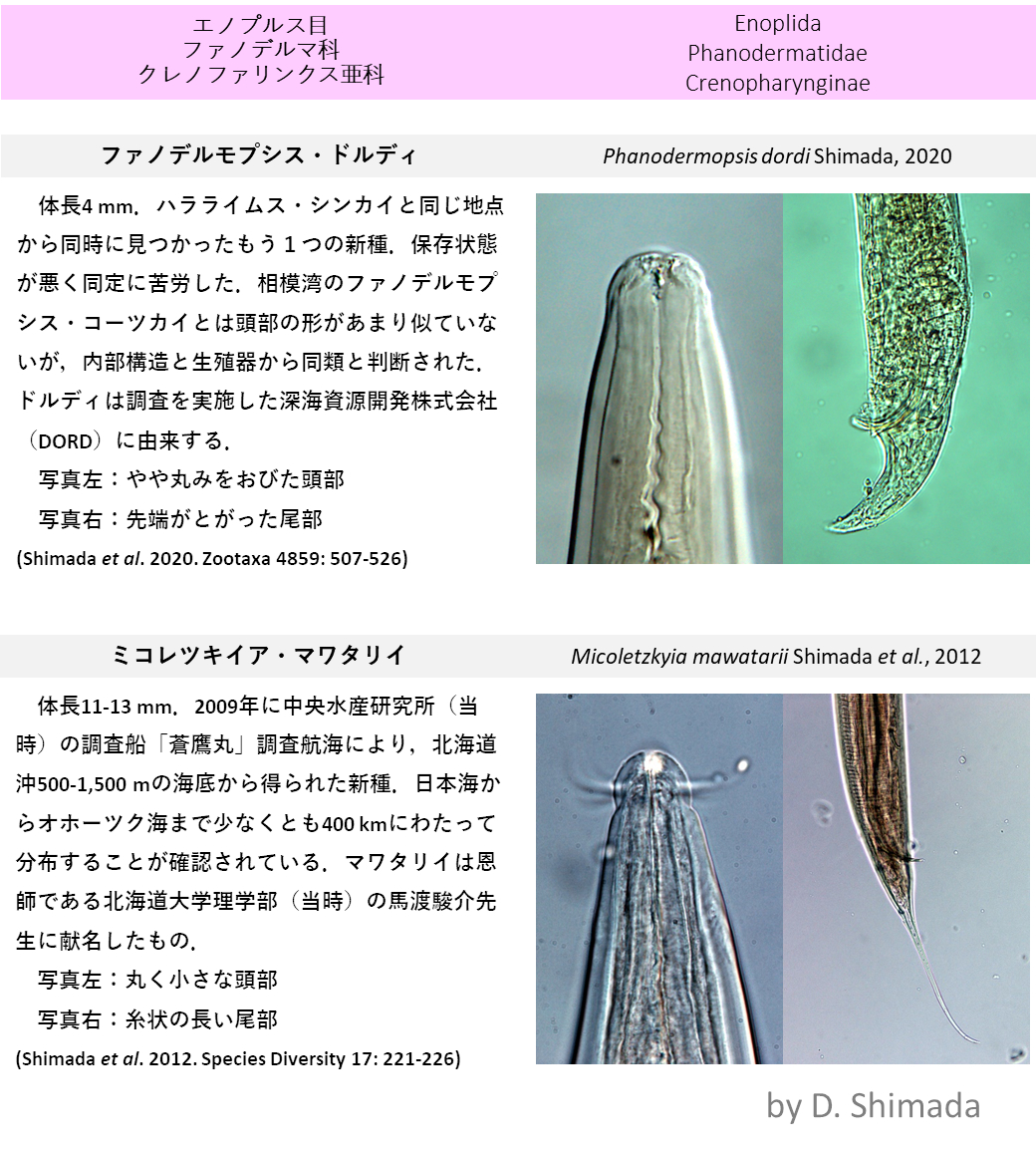
地球大気を簡略化した数値モデルにおいて、ジェット気流(左)から放射される大気重力波(中)の様子
ジェットの蛇行から生じる大気重力波の数値実験(球面浅水系)
大気重力波は浮力を復元力とする波である。重力波は下層から上空に伝播し、その運動エネルギーを中層大気(成層圏や中間圏)に供給し、この領域での大規模な大気の循環を駆動する。中層大気の大規模循環はオゾンや二酸化炭素の物質循環を通して、気候変動に大きな役割を担う。本研究では地球の自転効果の緯度変化を再現できる球面浅水系を用いて、ジェット気流の蛇行(左図:渦度場)とそれに伴う大気重力波の放射過程(中図:発散場)を、数値計算により調べている。右図は渦度場にバランスした水深であり、この系における高・低気圧渦の発生を表している。
大気重力波は浮力を復元力とする波である。重力波は下層から上空に伝播し、その運動エネルギーを中層大気(成層圏や中間圏)に供給し、この領域での大規模な大気の循環を駆動する。中層大気の大規模循環はオゾンや二酸化炭素の物質循環を通して、気候変動に大きな役割を担う。本研究では地球の自転効果の緯度変化を再現できる球面浅水系を用いて、ジェット気流の蛇行(左図:渦度場)とそれに伴う大気重力波の放射過程(中図:発散場)を、数値計算により調べている。右図は渦度場にバランスした水深であり、この系における高・低気圧渦の発生を表している。
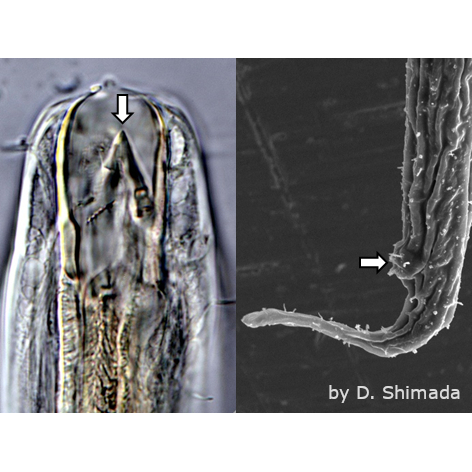
コケの葉の上を歩くオニクマムシ
Milnesium tardigradum

コケを住処とするクマムシは、活動状態の時には、体表に水の薄衣をまとっている。つまり、陸上種とはいえ、このようなクマムシは、間隙水の中で生活する「水生動物」なのである。環境が乾燥した状態では、昆虫のように活動することは不可能である。乾燥時はクリプトビオシスの状態で活動を休止する。これは「乾眠」とも呼ばれている。この能力を持つものだけが、コケを住処とすることができる。淡水産や海産のクマムシには、このような能力はみられない。それらは通常の動物と同様に、乾燥すると、「樽」ではなく「干物」になる。

磁場を印加しない場合の液体金属ダクト流れの乱流構造

車や飛行機の周りの空気の流れなど,身の回りには乱れた流れである乱流が存在する。その乱流中では大きなスケールの渦から小さなスケールの渦へ分裂していき,熱になって最後は散逸する過程が含まれている。上図では四方を壁で囲まれた矩形ダクト内を流体が右上から左下へ乱流状態で流れている様子を描画している。白い領域が強い回転渦が存在する領域で,流れ方向に渦が伸張している様子が分かる。赤と青の領域はそれぞれレイノルズ応力が正・負となる領域を示しており,その領域で乱流の生成や散逸が活発に起こっている様を表している。
スメクチック液晶相の偏光顕微鏡写真

液晶は流動性と配向秩序を併せ持つため、偏光顕微鏡を用いると、ドメインの欠陥を模様(テクスチャー)として観察できる。上の写真は、室温以下に融点を示し、結晶化しにくい有機塩(イオン液体)に、長鎖アルキル基を化学修飾したイオン性液晶のテクスチャーである。この液晶相では、アルキル基は結晶化、塩は融解しているため、高粘度でありながら高いイオン伝導度を示した。一般的に、イオン間には静電相互作用が強く働くため、塩は長鎖アルキル基よりも高温側で融解する。本研究で設計したイオン性液晶は、構成要素としたイオン液体の性質を反映したユニークな液晶相を発現した。
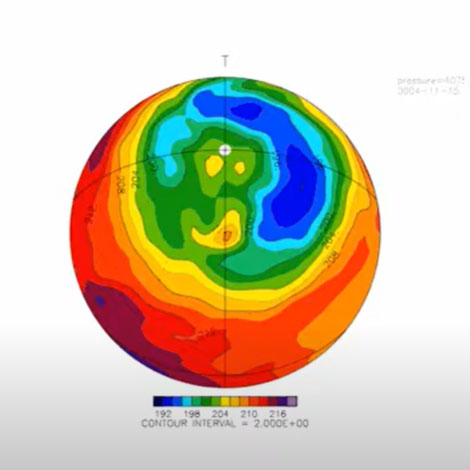
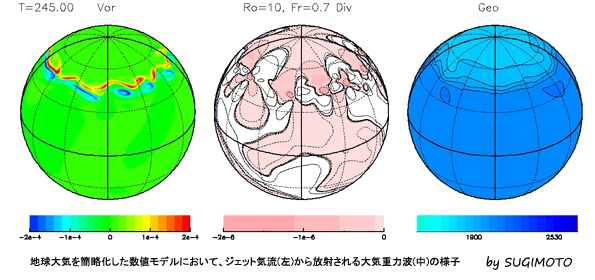
金星大気のコールドカラーの再現
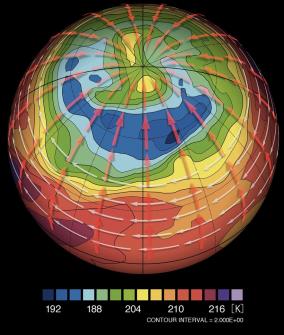
金星大気の雲層の上端では、暖かい極渦(緑、黄色)の周りを取り囲む「コールドカラー」と呼ばれる低温域(青)が存在する。我々のグループでは、金星大気大循環モデルを用いて、世界で初めてこの構造を再現した。南北風を赤の矢印、東西風を白の矢印で模式的に示している。


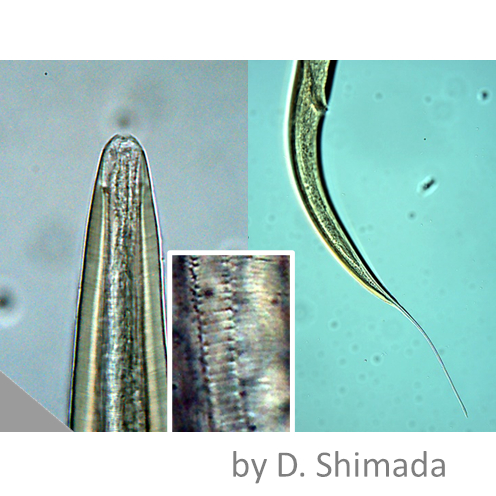
乾燥するにしたがって「樽」へ変身するオニクマムシ
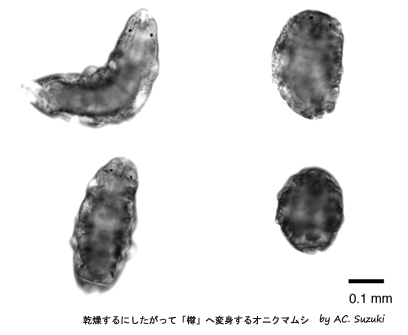
コケの葉の隙き間を住処とするクマムシは、環境が乾燥すると積極的に自ら乾燥し、樽のような形になる。これはクリプトビオシスと呼ばれる代謝停止状態である。代謝が停止した状態というのは、一般的には「死」を意味するが、クマムシ、センチュウ、ヒルガタワムシ等の動物は、その状態のまま生きながらえ、再び水分が補給されれば蘇って活動を再開する。「樽」となったクマムシは、酸素を必要とせず、絶対零度という低温や、100℃以上の高温、紫外線や放射線などに対する抵抗性を持つ。2007年に欧州で行われた宇宙実験では、地球軌道上で太陽光線に暴露されたオニクマムシの「樽」は、通常の生物ならば全滅する条件の中で、ごく少数ながら生き延びた、と報告されている。
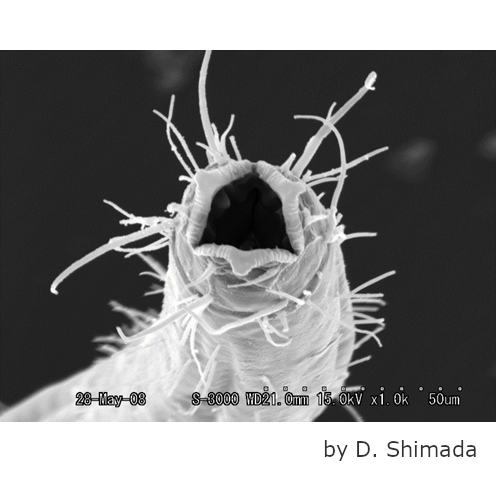
モミジガイ(
Astropecten scoparius)ビピンナリア幼生
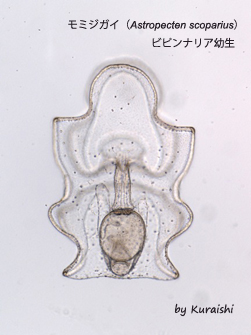
モミジガイは砂泥底の浅い海でよく見られるヒトデの仲間である。親は5放射の星形であるが、幼生は左右相称の形をしておりプランクトンとして海中を漂う。写真は受精後5日目のビピンナリア期の幼性で、体長は500µmほど。植物プランクトンを食べて成長する。
イトマキヒトデ(
Asterina pectinifera)のブラキオラリア幼生
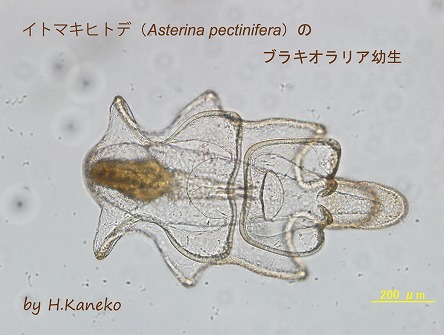
イトマキヒトデの発生過程において、原腸胚は口形成を完了すると、自ら餌を食べ、餌を消化できるビピンナリア幼生期に到達する。続いて、実験室の20℃の人工海水中で、植物性プランクトンを投餌して、ビピンナリア幼生を飼育すると、約1週間でこのような形態をしたブラキオラリア幼生に成長する。これ以後、さらに腕が伸長し、水槽中の小石などに反応して、稚ヒトデに変態する。ブラキオラリア幼生の身体は透明性が高く、食道や胃などの消化管の動きが容易に観察できる。この画像は、口と食道は収縮しており、まさに餌を胃へ送り込んでいる状態を示している。なお、帯状に肥厚した繊毛帯が身体の外壁を縁取りしていることも見て取れる。




イトマキヒトデ(
Asterina pectinifera)成体原基に形成される骨片
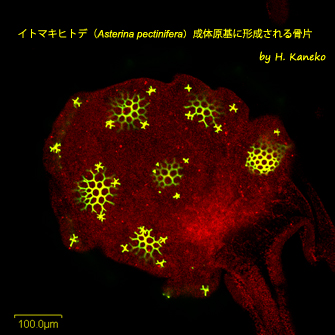
ウニと異なり、多くのヒトデ種は胚期に骨片を形成しない。しかしながら、ヒトデ種はブラキオラリア幼生期に到達すると、幼生体の後部に成体原基が生じ、変態後の成体ヒトデの固い殻をなす成体骨片を形成し始める。画像は、カルセインと呼ばれる物質を人工海水中に溶かし、イトマキヒトデのブラキオラリア幼生の飼育過程で、成体骨片を蛍光顕微鏡下で可視化したものである。ここでは、中央に1組、周囲に5組の成体骨片が観察されるが、成体原基の反対側にも少し形が異なった5組の骨片が形成され、計11組の骨片パターンとなる。これらの骨片がどのようにして形成されるのかは、まだヒトデ種では解明されていない。
モミジガイ(
Astropecten scoparius)後期ビピンナリア幼生

モミジガイの幼性は受精後2週間もするとからだの後部に成体の器官を作り始める。写真は受精後約3週間の幼性で、体長は1mmを超えている。イトマキヒトデの幼性と異なり、付着器であるブラキオラリア腕を作らないので、この時期でもビピンナリア幼生とよばれる。幼生の左側(写真は腹側から見ているので、向かって右側)には親の口側(基質に接する側)の構造が、右側には親の反口側(普通に観察した時に見える側)の構造ができる。このため幼生後部に限ってみると、左右の不相称性が際立っている。